旭月的售后服务还是非常不错的,不论是工作日还是周末的问题咨询,工程师都能在第一时间给予解答,从而保证我们的实验可以顺利进行;联盟微信号对最新科研成果的推送,以及每年免费整理赠送的论文集,可以让用户了解非损伤微测技术的应用领域并掌握自己研究领域的最新动态。这些举措都对非损伤微测技术的广泛使用具有重要的推动作用,相信在联盟和旭月的带领下,非损伤微测可以越走越远。给巨工点赞,给旭月点赞,给联盟点赞!
我认为抗盐研究要重视NMT,围绕NMT可以做很多工作,不能只看做是锦上添花的数据。对于研究生,把NMT这类技术学精,将是他们学习生涯的关键收获。
从仪器安装至今一年半期间,无论是仪器设备的操作还是实验过程中遇到的各种问题,旭月的工程师总能给予及时专业的帮助。购买耗材也经常有优惠券,非常棒!NMT方面的研究项目和相关文献汇总的非常及时,对我们很有参考价值。
非损伤微测技术为植物营养研究提供了一个新的、可靠的技术手段,增加了研究的创新性。相比其他同类型的公司,旭月公司带领的非损伤微测技术团队也非常专业。我们利用该技术对农作物营养、非生物胁迫方面等进行了多项研究,申请了多个国家自然科学基金,帮助我们在离子通道、离子跨膜转运/吸收等方面有了很大突破。
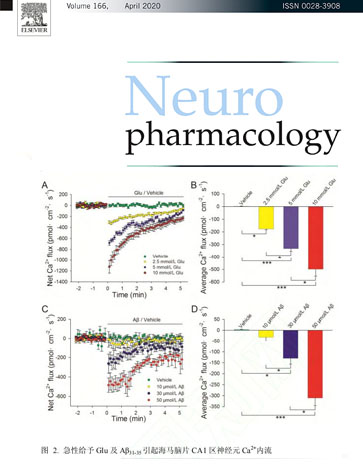
非损伤检测系统能够帮助我们非常直观、无损及量化地看到各种离 子在乳腺癌组织或细胞 内外浓度的变化情况,从而可以判断出离子吸收 或排出的情况,是一种研究乳腺癌微环境非常好的技术手段。近些年,我们利用旭月公司的非损伤微测技术在乳腺癌肿瘤组织生 理特征及其乳腺癌治疗中的潜在应用进行了富有成效的有益探索。这些 探索还处于初始阶段,但在世界范围内是具有开拓意义的。
作为旭月公司非损伤微测技术的十多年的老用户,我们 利用该技术在耳鼻喉基础研究领域,进行了有益的研究和探 索。同时也见证了旭月公司从引进、消化、吸收到再创新、 并达到世界领先水平的整个历程。
非损伤微测技术是具有世界先进水平的多学科 (微 电子、计算机、精密机械加工、物理、数学、高分子化学、碳纳米及光学显微技术)尖端科技的系统集成。该技术平台在在电脑自动控制下,在不接触被测样品的情况下三维、实时、动态地获得进出样品的各种分子和离子浓度、流速及其运动方向的信息,获得其他技术难以测到的生理特征和生命活动规律。旭月公司将世界领先水平的非损伤微测技术引进国内,发扬光大,促进了我国在该技术领域的发展。
非损伤微测技术可在不损伤样品的前提下检测分子/离子进出生物活体的流速(运动速率和方向),可以作为研究水稻养分吸收和利用的有利工具。利用非损伤微测技术从水稻根系的铵冗余吸收、铵钾离子吸收互作等角度解析品种间、环境间氮肥利用差异,拓展水稻养分利用研究的视野;在该技术的帮助下,我成功申请到2个国家自然科学基金面上项目资助,并于2020年被评为浙江省“万人计划”创新领军人才。
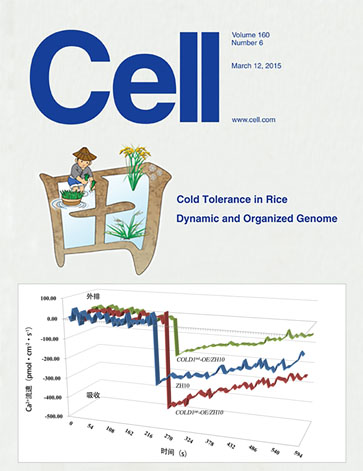
作为旭月公司非损伤微测技术的十多年的老用户、老朋 友,我们利用该技术在水稻栽培生理等方面进行了卓有成效 的研究和探索,相 关研究成果发表于 《The Plant JOurna1》 《Plant Cell and Environment》 等国内外著名学术期刊。同时 也见证了旭月公司从引进、消化、吸收到再创新、并达到世 界领先水平的整个历程。
离子/分子跨膜运输活性的直接检测对阐述植物逆境适应机制,揭示重要基因的生物学功能至关重要,NMT在离子/分子运输活性检测方面具有显著优势,对本课题组研究工作的开展具有较大促进作用。利用NMT,本课题组在植物离子平衡调控机制和功能基因挖掘方面取得了一些突破和进展,在Plant Phyology、Journal of Experimental Botany、Horticulture Research等重要植物学期刊发表多篇研究论文。
非损伤微测系统能够帮助我们非常直观、无损地看到金属离子、盐离子在植物根系或细胞内外浓度的变化情况,从而可以判断出离子吸收或排出的情况,为林木育种提供证据。中国林科院亚热带林业研究所于2014年购置了双通道非损伤微测系统,迄今为止,先后开展了林木耐盐、植物重金属富集以及转基因材料的胁迫生理分析,在该系统的支持下,先后在Environmental Science&Technology、The Plant Biotechnology Journal、Tree Physiology、Chemosphere等SCI刊物上发表了一系列论文,筛选出植物重金属富集新基因5个,耐盐转基因杨树10个,重金属转基因杨树5个。
拥有5年以上非损伤微测系统操作应用、售后经验,累计售后服务近三十家单位,售后服务经验2000+小时。
美国扬格(旭月北京)售后工作负责人,拥有6年以上非损伤微测系统操作应用、售后经验,累计售后服务近六十家单位,售后服务经验3500+小时。Cu、Pb流速传感器商业化研发团队骨干成员。
服务过近百位测试客户,具有丰富的测试经验,多次委派至创服中心进行测试工作。
拥有10年以上的非损伤微测系统的使用经验,服务过上百位的测试客户,是IAA传感器研发团队的骨干成员。
美国扬格(旭月北京)中国华东区售后负责人,高级培训工程师,拥有近3年的售后服务经验,累计服务近三十家单位,售后服务经验1300+小时。





 |
 |
 |
 |
 |
 |
总公司地址:北京市海淀区彩和坊路10号1号楼(1+1大厦)8层808
电话:010-8262 2628
分公司地址:江苏省常州市钟楼区长江中路299号 中博创业园4幢126室